1910 ÉS 1919 KÖZÖTT egy amerikai genetikus, THOMAS HUNT MORGAN felismerte, hogy a Mendel által megállapított 9:3:3:1 aránnyal ellentétben hasadási aránytól, számos jelleg hajlamos a kapcsolt öröklődésre. Ennek a génkapcsolódásnak a legnyilvánvalóbb példája a nemi kromoszómákkal összefüggő jellegek öröklődése. Az emberben és számos más gerincesben is a nőnek két X kromoszómája, a férfinek pedig egy X és egy Y kromoszómája van. (1991-ben londoni kutatók lokalizálták az Y kromoszóma azon génjét, amely a hímneműségért felelős). A nemi (szex-) kromoszómák ezen kívül más jellegek génjeit is hordozzák, amelyeket ennek megfelelően nemhez kapcsolt géneknek neveznek. Az emberek vörös-zöld színvakságát az X kromoszómán levő recesszív gén határozza meg. A gént ugyan a nő is hordozza, de ritkán szenved hatásától, mivel gyakran elfedi azt a másik X kromoszómán levő domináns gén, amely a normális színlátást teszi lehetővé. Ez a betegség tehát gyakoribb a férfiaknál.
Ecetmuslicákkal dolgozva, MORGAN úgy találta, hogy még az azonos kromoszómán elhelyezkedő jellegek is újrarendeződhetnek egymás között. Annak ellenére, hogy ez nem történik meg túl gyakran, de MORGAN gyanítani kezdte, hogy a kromoszómák a meiózis (és mitózis alatt [is]) eltörhetnek és azután rekombinálódnak.
Ezután arra gondolt, hogy két, egymástól a kromoszómán távol levő gén között a törésnek az esélye nagyobb mint két olyan gén között, amelyek a kromoszómán egymáshoz közel helyezkednek el. Ha ez így van, akkor az a gyakoriság, amivel a gének kicserélődnek egymás közt, felhasználható annak a meghatározására, hogy az adott gének egymástól milyen távolságra helyezkednek el a kromoszómán. Olyan ecetmuslicákat keresztezve, amelyek sok különböző jelleget hordoztak, MORGAN bebizonyította, hogy igaza volt. A végeredmény kromoszómatérképek sorozata volt, amely a gének lineáris elhelyezkedését mutatta az ecetmuslica kromoszómáin. A kutatók ma ugyanezen az elven alapuló módszerrel, sokkal precízebben készítik el az emberi és más kromoszómák térképeit. Új génkombináció kialakulása a legtisztábban az ivarsejtképződés során látható. Először is a kromoszómapárok egymáshoz közel kerülnek a meiózis alatt. Mindegyik kromoszóma két kromatidára válik szét és ezek anyaga véletlenszerűen cserélődik ki az apai és anyai kromoszómák között. Mutációk (beleértve a genetikai anyag kitörlődését is) előfordulhatnak az átkeresztezés (crossing over) alatt. Második lépésben a kromoszómák a meiózis alatt újrarendeződnek, mindegyik gaméta egy sorozat véletlenszerűen összerendeződött anyai és apai kromoszómából álló sorozatot kap. Még ha csak négy kromoszómáról van is szó, akkor is 16 különböző kombináció lehetséges már a gamétákban, és ez 16 x 16 = 256 kombinációra ad lehetőséget a két gamétából keletkező utódban.
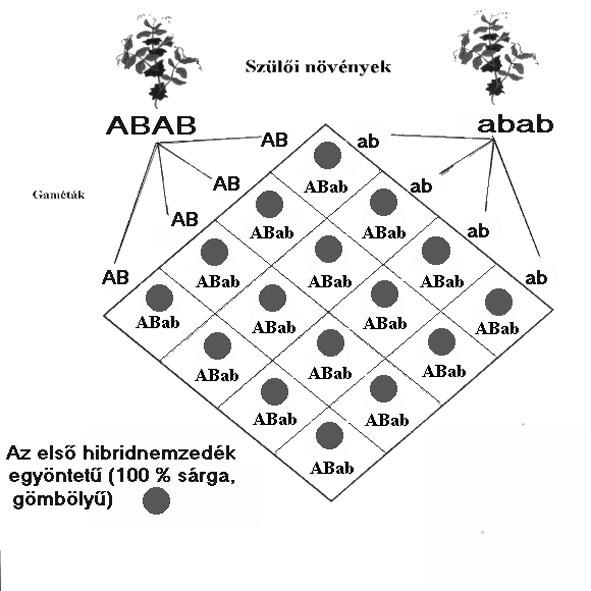
I. NEMZEDÉK
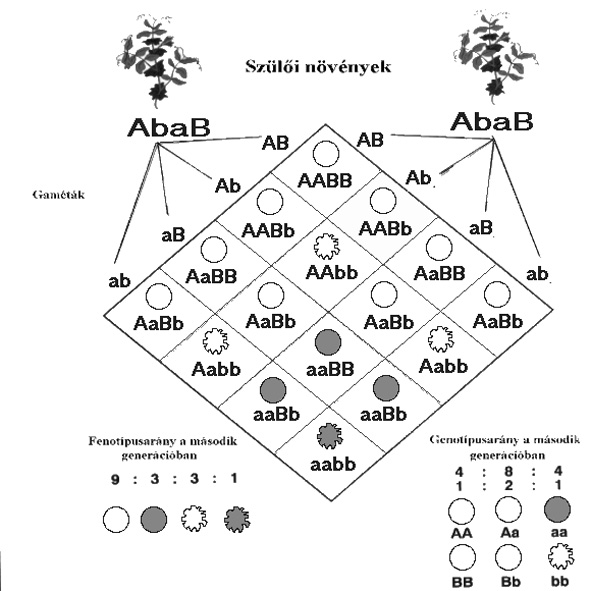
II. NEMZEDÉK
Negyven évvel ezelőtt, az 1953 májusában és áprilisében kiadott publikációkban az angol fizikus, FRANCIS CRICK és az amerikai biológus JAMES WATSON bebizonyította, hogy a DNS kettős hélixet alkot. A molekulák központjában bázisok állnak:
A bázisok közötti kötések tartják össze a két hélixet. Az A mindig a T-nel kapcsolódik, a C mindig G-nal. WATSON és CRICK kettős hélixe világosan megmagyarázta, hogy a genetikai információ hogyan másolódik egyik sejtből vagy egyik nemzedékből a másikba. Az egymást kiegészítő fonalak kicsavarodnak és az új partner felépítéséhez szükséges építőelemek mindkét szálhoz odaszállítódnak. A bázisok sorrendje a DNS-ben meghatározza a fehérjék felépítésében résztvevő aminosavakat, éppúgy, mint ahogyan a morzejelek pontjai és vonalai az ábécé betűit jelzik. Egy adott gén - a DNS szegmense (részlete) - meghatározza egy adott fehérje (vagy fehérje-részlet) megfelelő szerkezetét. Minden aminosavat három bázis (egy kodon) határoz meg. Négy különböző bázis van; ha egyszerre hármat veszünk, akkor ezek 4 x 4 x 4 = 64 lehetséges kodont kódolhatnak. Ez több mint elégséges ahhoz, hogy a fehérjékben megtalálható 20 aminosavat meghatározzák. A threonint például nemcsak az ACT határozza meg, hanem éppúgy az ACC, ACA és ACG kombinációk is.
| U | C | A | G | ||
| U | PHE | SER | TYR | CYS | U |
| U | PHE | SER | TYR | CYS | C |
| U | LEU | SER | stop | stop | A |
| U | LEU | SER | stop | TRP | G |
| C | LEU | PRO | HIS | ARG | U |
| C | LEU | PRO | HIS | ARG | C |
| C | LEU | PRO | GLN | ARG | A |
| C | LEU | PRO | GLN | ARG | G |
| A | ILE | THR | ASN | SER | U |
| A | ILE | THR | ASN | SER | C |
| A | ILE | THR | LYS | ARG | A |
| A | MET | THR | LYS | ARG | G |
| G | VAL | ALA | ASP | GLY | U |
| G | VAL | ALA | ASP | GLY | C |
| G | VAL | ALA | GLU | GLY | A |
| G | VAL | ALA | GLU | GLY | G |
A DNS ezenkívül tartalmaz olyan géneket is, amelyek
a fehérjeszintetizáló géneket a szükségleteknek megfelelően ki- illetve bekapcsolják.
A baktériumok pl. így válaszolnak a környezet
változásaira. Az állati sejteknek azonban állandó környezetük van, amelyet más
szövetek és testnedvek biztosítanak. Ezért, annak ellenére, hogy a különböző
szövetek - mint a bőr-, izom- és idegszövet - sejtjei ugyanazt a DNS-t tartalmazzák
kromoszómáikban, ezekben a sejtekben csupán azok a gének képeznek enzimeket
és más fehérjéket, amelyek a szövet specializált működéséhez szükségesek. A
megtermékenyített petesejtből származó differenciálódott sejtek génjei nem vesznek
el, csupán az történik, hogy hatásuk nem érvényesül.
Egy bőrsejt nem kezd el úgy működni mint egy májsejt, és viszont.
